Treatment of ethylnitrosourea induced lymphocyte hyperproliferation by DNA hypomethylation in the
rat colon
ÖZDEMİR, Öztürk
Department of Medical Biology and Genetics, Faculty of Medicine, Cumhuriyet University, 58140-SIVAS, TURKEY
BULUT, Hüseyin Eray
Department of Histology - Embryology, Faculty of
Medicine, Cumhuriyet University, 58140-SIVAS, TURKEY
KORKMAZ, Mehmet
Department of Medical Biology, Health High School, The University of Balıkesir, BALIKESIR, TURKEY
EĞİLMEZ, Reyhan
Department of Pathology,
Faculty of Medicine, Cumhuriyet University, 58140-SIVAS, TURKEY
ATALAY, Atilla
Department of Biochemistry, Faculty of Medicine, Cumhuriyet University, 58140-SIVAS, TURKEY
Correspondence
should be addressed to: ÖZDEMİR, Öztürk, PhD.
Department of Medical Biology and Genetics, Faculty of Medicine, Cumhuriyet University, 58140-SIVAS, TURKEY
Tel : + 90 346 219 10 10 /
1068 Fax: + 90 346 219 11 55
Email: ozdemiro@bim.cumhuriyet.edu.tr
Submitted for
publication: 12-05-2000
Keywords: :
5-aza-2'-deoxycytidine, antineoplastic effect, colon, rat



ABSTRACT
N-ethyl-N-nitrosourea (ENU) is
a potential carcinogenic agent commonly used in industry, and it may cause an
uncontrollable cell proliferation and eventually tumourgenesis. On the other
hand, the hypomethilation of DNA by 5-aza-2'-deoxycytidine is the best known
anti-tumoural mechanism used for the treatment of leukemia. Therefore the
present study aimed to find out the possible healing effects of 5-aza-2'-deoxycytidine
on lymphocyte hyperproliferation in the rat colon through the above mentioned
DNA hypomethylation mechanism.
Rats were injected with 300mg/kg body weight - body
weight ENU (i.p.) in order to induce tumour development. Following 45
weeks when the tumourgenesis was proved visually, animals were treated with
5-aza-2'-deoxycytidine 100mg/100 g body weight twice a week intraperitoneally for
15 weeks. After the experimental procedure, all animals were sacrificed and
colonal tissues were obtained. Tissues were processed for light and electron
microscopy.
While no
colonal tumour development was observed in the control group, an extensive
tumour development was seen in the subcutaneous region in the high dose ENU
treated group. The light and electron microscopical examination of the rat
colonal tissue revealed a lymphocyte hyperproliferation and invasion in the
submucosal region, an increased number of polimorphonuclear leukocytes (PMNLs)
and occasional epithelial lesions. On the other hand, the evaluation of the
5-aza-2'-deoxycytidine treatment group rat colon demonstrated features similar
to those seen in the control and PEG treated groups indicating a possible
anti-neoplastic effect of 5-aza-2'-deoxycytidine via DNA hypomethylation.
INTRODUCTION
It has been shown that ENU
induces tumour development in various organs of mammalian species (1, 2). This monofunctional alkylating agent
is a potent inducer of cellular stress leading to chromosomal abberations such
as point mutations, translocations, deletions, insertions and cell killing (3, 4). This agent is also known as a potent
cell mutagen due to its alkalating function and induced DNA damage in the cell
(5 -7).
5-aza-2'-deoxycytidine (5-azadCR, DAC, Decitabine) and
its ribose congener 5-azacytidine are pyrimidine analogs and specific
inhibitors of DNA methyltransferase enzyme. Both drugs are able to reduce the
biochemical activity of DNA methyltransferase enzyme in cells. A great deal of
preclinical studies have shown that 5-azadCR is able to induce maturation of
human leukemic cells and inhibit clonogenic potential of cells in vitro in the absence of acute cell
killing with a mechanism involving DNA hypomethylation (8).
These cytidine analogs, modified in position 5, were originally developed as
antitumour drugs, and have been used in the treatment of both childhood and
adult leukemias (9).
The DNA methyl inhibitor, 5-azadCR, also causes
chromosome decondensation in the human lymphocyte cell culture (10).
5-azadCR is the most potent specific inhibitor of DNA methylation (11). Agent was demonstrated to be an active antileukemic drug (12, 13). Pinto et al presented
evidence for in vivo induction of
leukemic cell differentiation by cytidine analogs (14).
The present study aimed to investigate the effects of
5-azadCR treatment on hyperprolipherated lymphocyte cells of rat colon that
induced by a potent carcinogen and an alkylating agent, ENU.
MATERIALS AND
METHODS
ANIMALS
Animals used in the present
study were non - transgenic and were bred and fed in optimal laboratory
conditions. Fourty male 7-8-week old wistar albino rats (Rattus norvegicus),
obtained from "The Experimental Animal Laboratory of Cumhuriyet
University, Sivas - Turkey", were used in the present study. Three group rats
were used in the present study; control, ENU treatment and
5-aza-2'-deoxycytidine treatment groups.
EXPERIMENTAL DESIGN
N'-ethyl-N'-nitrosourea (ENU)
(CASRN : 759-73-9, Sigma Chemical Company, MO, USA) and polyethylene glycol
(PEG) were obtained from Sigma Chemical Company, USA. ENU is dissolved in PEG
and stored at -700C. While the experimental group rats (15 male and
15 female in a total of 30) were injected once a week with 300mg/kg - body
weight with ENU (i.p.), and the control group animals received no agent administration.
Following 45 weeks of experimental period, 10 rats per treatment group were
sacrificed and colonal tissues were obtained. The rest of the animals (20 rats)
from ENU treated group were used for 5-azadCR administration.
5-AZA-2'-DEOXYCYTIDINE
(5-AZAdCR ) ADMINISTRATION
5-azadCR was purchased from
Sigma Chemical Co. Stock solutions of drug were prepared in phosphate-buffered
saline (PBS) at a concentration of 25mg / 2ml (55 mM), aliquoted and stored at
-40 0C. Each aliquot was used once only, thawed immediately before
use, serially diluted with distilled water, and treatment group rats were
injected intraperitoneally in 100mg/100 g body weight or 0.5
mM
of final concentration for 2 time a week (approximately 72 hours). Following 15
weeks of drug administration, all animals were killed and colonal tissues were
obtained.
MICROSCOPY
For light microscopy, colonal
tissue samples were fixed in 3% glutaraldeyde, dehydrated through the
increasing concentrations of ethanol and embedded in JB4 glycol metacrylat. 2 -
2.5 mm
thick sections were stained with toluidin acid fuchsin - blue. They were
evaluated under a Jenamed 2 (Carl Zeiss Jena, Germany) light microscope, and
appropriate field of views were photographed.
Tissue samples for electron microscopy were fixed with 3%
glutaraldehyde in 0.1M phosphate buffer (pH 7.4) for 2-4 hours. Tissues were
rinsed with buffer, post-fixed in 1 % aqueous osmium tetroxide for 2 hours,
dehydrated in ethanol, and embedded in Epon resin. Semithin tissue sections
were stained with toludin blue and evaluted at the light microscopical level
wheareas the ultra-thin sections were double stained with uranyl acetate - lead
citrate, and observations were done at the ultrastructural level using a Zeiss
(Germany) electron microscope.
RESULTS
All ENU treated rats exhibited
large subcutaneous tumours (approximately 5 - 9 cm in size) at different body
regions when evaluated visually. These tumours were also investigated
histopathologically and showed distinct characteristics of subcutaneous
gliosarcoma (data not shown). Control group rat colonal tissues demonstrated
mucosal crypts that contained simple columnar epithelium and underlying lamina
propria in their regular appearances (Figure 1).
{kind=link}
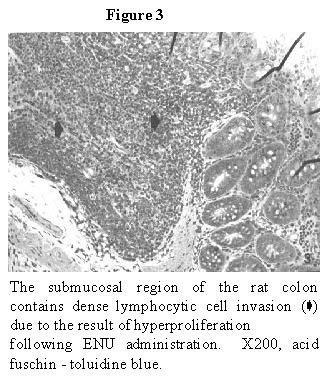
The ENU treated group rat colonal features revealed
distinct histopathological changes such as mucosal and submucosal
hyperproliferation of lymphocyte cells and lymphocytic invasion into the
mucosal layer from the underlying submucosa (Figure 2
and Figure 3). These histopathological alterations
must be the result of ENU treatment since the PEG - only administration caused
no such changes (data not shown). Electron microscopical examination of the ENU
treated group submucosal region of the rat colon supported the
hyperproliferation of lymphocytes (Figure 4).
{kind=link}
{kind=link}
{kind=link}
After the 5-azadCR administration of the ENU treated
rats, it was seen that the drug reduced the number of lymphocytes in colonal
tissues (Figure 5a, b). The electron microscopical
examination of 5-azadCR treated rat colon revealed ultrastructural features
similar to the control group (Figure 6). However,
an increased protein content was present in the 5-azadCR treated group stromal
tissue possibly due to DNA hypomethylation and distinc gene reactivation (Figure 7).
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
A consistent amount of data
has been accumulated over the last 15 years about the clinical activity and
biological properties of 5-azadCR in leukemic diseases. The results of in vitro and in vivo studies showed that the drug is a powerful antileukemic
agent and displays healing activity in AML and ALL patients. As a reversible
epigenetic modification which can affect gene expression and DNA
hypomethylation has been attractive candidate for the biochemical mechanism of
genomic imprinting (15). Transcriptional blocks in p16INK4A
and p15INK4B genes in gastric carcinomas were reversed by 5-azadCR treatment (16). Frequent aberrant methylation patterns of p16INK 4A gene
was reported in primary rat lung tumours (17).
Differential repair of structurally distinct mutagenic
lesions in particular genes may influence the cellular risk of malignant
conversion. Complete carcinogens must possess both initiating and promoting
properties in tumour development (18). While most N-nitroso
compounds are potential mutagens and considered to be tumour initiating agents,
some are not mutagenic and yet are complete carcinogens. The present study
investigated the ethylnitrosourea-induced rat colonal structural changes at the
light and electron microscopical levels and these structural changes were
treated with 5-azadCR which causes gene reactivation/DNA hypomethylation
possibly in the tumour suppressor p53 gene or in the related genes. The DNA
methyl inhibitor, 5-azadCR, took place significant role in the treatment of
hyperproliferative lymphocytes of rat colon in the present study. There were
neither tumours nor colonal tissue alterations in the untreated group whereas
distinct colonal lymphocyte hyperproliferation, invasion and lymphoepithelial
lesions were evident in the ENU treated rats. Sequential intra-peritoneal injections
of ENU (300 mg/kg) strongly induced subcutaneous sarcomas (data not shown) and
colonal tissue changes 45 weeks after treatment. In colon, however, ENU at its
highest dose caused adverse alterations in rat colonal tissue. Favor suggested
the threshold model for explaining the ENU mutagenity in germ cells claiming
that the doses below the threshold dose result in induced DNA adducts that are
repaired (19). ENU may possibly cause some changes in A/T to
G/C or G/C to A/T sequences (19, 20)
indicating that the high dose of ENU was effective in inducing mutations in
colonal tissue changes. Similarly the present study used a high dose of ENU and
found reversible alterations in the colon. Loss of the wild-type allele results
in a mutator phenotype, accelerating tumorgenesis which specifically occurs in
the gastrointestinal and genitourinary tracts (21,22).
In addition, the long term ENU exposure to rats may cause
different type of tumours at different organs, one of which was a very large
subcutaneous sarcoma that observed visually in rats 45 weeks after ENU
treatment in the present work. There is wide variation of AGT levels between
the organ and cell types, which appears to correlate with cell and tissue type
sensitivity to the mutagenic and carcinogenic effects of alkylating agents.
Findings of the present study supported the idea that the lymphocyte
hyperproliferation caused by ENU may possibly lead to the development of
lymphocarcinoma in the rat colon.
It could be postulated that ENU presumably initiates the
triggering signal for colonal carcinogenesis by alkylating the bases of
A/T-G/C, A/T-C/G, A/T-T/A, G/C-C/G and G/C-T/A base substitutions as suggested
by Shubiya (23,24). The injection of
5-azadCR reversed the rat colonal tissue changes caused by ENU due to DNA
hypomethylation. Therefore the 5-azadCR administration produced an
antineoplastic effect on the colonal lymphocyte hiperproliferation. In
addition, 5-azadCR treatment caused increased protein synthesis. In conclusion,
it could be postulated that 5-azadCR has the potential of tumour supressor gene
activation which have been spontaneously silenced by DNA hypermethylation in
the hyperproliferated lymphocytes.
CONCLUSIONS
It can be suggested that a distinct subcutaneous tumour development and a lymphocyte hyperproliferation which might be the triggering signal for carcinogenesis were observed in rat colon due to the alkylating characteristics of ENU. On the other hand, 5-azadCR treatment reversed those colonal alterations through the DNA hypomethilation mechanism. Although the molecular basis of this mechanism have not been completely explained, the altered methylation patterns and reactivation of some distinct genes could be an important step in tumour therapy.
ACKNOWLEDGEMENTS
Authors would like to thank to
" The State Planing Organisation of TURKEY (DPT/ 96 K120630) " for
funding this study and also thank to Professor M. KAYA and Professor S.POLAT of
Cukurova University for their technical support.
REFERENCES
1. Henderson L, Wolfreys A, Fedyk J, Bourner C, Windebank S (1998) The
ability of the Comet assay to discriminate between genotoxins and cytotoxins.
Mutagenesis 13(1): 89-94
2. Sasaki YF, Tsuda S, Izumiyama F, Nishidate E (1997)Detection of
chemically induced DNA lesions in multiple mouse organs (liver, lung, spleen,
kidney, and bone marrow) using the alkaline single cell gel electrophoresis
(Comet) assay. Mutat Res 15, 388(1): 33-44
3. Nikolova T, Huttner E (1996) Adaptive and synergistic effects of a
low-dose ENU pre-treatment on the frequency of chromosomal aberrations induced
by a challenge dose of ENU in human peripheral blood lymphocytes in vitro.
Mutat Res 25, 357(1-2): 131-141
4. Wilhelm D, Bender K, Knebel A, Angel P (1997) The level of
intracellular glutathione is a key regulator for the induction of
stress-activated signal transduction pathways including Jun N-terminal protein
kinases and p38 kinase by alkylating agents. Mol Cell Biol 17(8): 4792-4800
5. Op-het-Veld CW, van-Hees-Stuivenberg S, van-Zeeland AA, Jansen JG
(1997) Effect of nucleotide excision repair on hprt gene mutations in rodent
cells exposed to DNA ethylating agents. Mutagenesis 12(6): 417-424
6. Suzuki T, Hayashi M, Wang X, Yamamoto K, Ono T, Myhr BC, Sofuni T
(1997) A comparison of the genotoxicity of ethylnitrosourea and ethyl
methanesulfonate in lacZ transgenic mice (Muta Mouse). Mutat Res 5;
395(1):75-82
7. Tong HH, Park JH, Brady T, Weghorst CM, D'Ambrosis SM (1997)
Molecular characterization of mutations in the hprt gene of normal human skin
keratinocytes treated with N-ethyl-N-nitrosourea: influence of O6-alkylguanine
alkyltransferase. Environ Mol Mutagen 29(2): 168-179
8. Mandelli F (1993) Introduction to the workshop on DNA
methyltransferase inhibitors. Leukaemia, Suppl Monograph 1, 7:1-2
9. Taylor SM (1993) 5-aza2'-deoxycytidine: Cell differentiation and DNA
methylation. Leukaemia, Suppl Monograph 1, 7:3-8
10. Özdemir Ö, Sezgin İ, Çolak A (1998) Underorganisation of chromatin
structure in human metaphase chromosomes by induction of 5-aza-2'-deoxycytidine
(Decitabine). Gülhane Tıp Dergisi 40(4):420-427
11. Jones PA, Taylor SM (1980) Cellular differentiation, cytidine
analogs and DNA methylation. Cell 20:85-93
12. Momparler RL, Rivard GE, Gyger M (1985) Clinical trial on
5-aza2'-deoxycytidine in patients with acute leukaemia. Leukaemia Res
30:277-286
13. Rivard GE, Momparler RL, Demers J, Benoit P, Raymond R, Lin KT,
Momparler LF (1981) Phase I study on 5-aza2'-deoxycytidine in children with
acute leukaemia. Leukaemia Res 5:453-462
14. Pinto A, Zagonel V, Gattaei V, Marotta G, Bullian PL, Mancardi S,
Coglievina M, De Rosa L, Alosi M, Carbone A, Attadia V (1990) 5-aza2'-deoxycytidine
as a differentiation inducer in human hemopoietic malignancies: preliminary
observations on the in vivo modulation of leukaemia cell phenotype and
correlation with clinical response. In: preclinical and clinical studies.
Haarlem, The Netherlands: PCH Publications, p 143-164
15. Tycko B (1997) DNA methylation in genomic imprinting. Mutat Res
386(2):131-140
16. Lee YY, Kang SH, Seo JY, Jung CW, Lee KU, Choe KJ, Kim BK, Kim NK,
Koeffler HP, Bang YJ (1997) Alterations of p16INK4A and p15INK4B genes in
gastric carcinomas. Cancer 80(10): 1889-1896
17. Swafford DS, Middleton SK, Palmisano WA, Nikula KJ, Tesfaigzi J,
Baylin SB, Herman JG, Belinsky SA (1997) Frequent aberrant methylation of
p16INK4A in primary rat lung tumors. Mol Cell Biol 17(3):1366-1374
18. Brustle O, Petersen I, Radner-H; Holl T, Plate KH, Kleihues P,
Wiestler OD (1993) 1993; Complementary tumour induction in neural grafts
exposed to N-ethyl-N-nitrosourea and an activated myc gene. Carcinogenesis
14(8): 1715-1718
19. Favor J (1998) The mutagenic activity of ethylnitrosourea at low
doses in spermatogonia of the mouse as assessed by the specific-locus test.
Mutation Research 405: 221-226
20. Cerutti P, Hussain P, Pourzand C, Aguilar F (1994) Mutagenesis of
the H-ras protooncogene and the p53 tumour suppressor gene. Cancer Res 1, 54 (7
Suppl): 1934 -1938
21. de-Wind N, Dekker M, van-Rossum A, van-der-Valk M, te-Riele H
(1998) Mouse models for hereditary nonpolyposis colorectal cancer. Cancer Res
15, 58(2): 248-255
22. Shoemaker AR, Luongo C, Moser AR, Marton LJ, Dove WF (1997) Somatic
mutational mechanisms involved in intestinal tumor formation in Min mice.
Cancer Res 15; 57(10): 1999-2006
23. Pourzand C, Cerutti P(1993) Genotypic mutation analysis by
RFLP/PCR. Mutat Res 288(1):113-121
24. Shibuya T, Morimoto K (1993) A review of the genotoxicity of
1-ethyl-1-nitrosourea. Mutat Res 297(1): 3-38